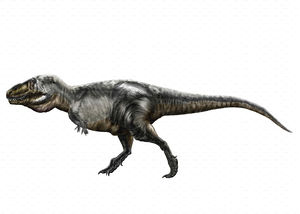
 Тиранноза́вр[1] (Шаблон:Lang-la — «ящер-тиран», от Шаблон:Lang-grc [tyrannos] «тиран» и Шаблон:Lang-grc2 [sauros] «ящер, ящерица»[1], также встречается неправильное написание тиранозавр[1]) — монотипный род плотоядных динозавров из группы целурозавров, подотряда тероподов, включающий единственный вид — Tyrannosaurus rex (Шаблон:Lang-lat «царь»).
Тиранноза́вр[1] (Шаблон:Lang-la — «ящер-тиран», от Шаблон:Lang-grc [tyrannos] «тиран» и Шаблон:Lang-grc2 [sauros] «ящер, ящерица»[1], также встречается неправильное написание тиранозавр[1]) — монотипный род плотоядных динозавров из группы целурозавров, подотряда тероподов, включающий единственный вид — Tyrannosaurus rex (Шаблон:Lang-lat «царь»).

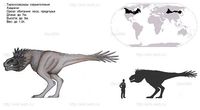
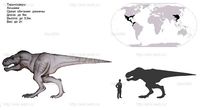 Обитал в западной части Северной Америки, которая в те времена представляла собой остров Ларамидию, и был наиболее распространённым из тираннозавридов. Окаменелые останки тираннозавров находят в различных геологических формациях, датирующихся маастрихтским ярусом позднемелового периода, около 67—65,5 миллионов лет назад[1]. Был одним из последних ящеротазовых динозавров, существовавших перед катаклизмом, положившим конец эре динозавров (мел-палеогеновым вымиранием).
Обитал в западной части Северной Америки, которая в те времена представляла собой остров Ларамидию, и был наиболее распространённым из тираннозавридов. Окаменелые останки тираннозавров находят в различных геологических формациях, датирующихся маастрихтским ярусом позднемелового периода, около 67—65,5 миллионов лет назад[1]. Был одним из последних ящеротазовых динозавров, существовавших перед катаклизмом, положившим конец эре динозавров (мел-палеогеновым вымиранием).
Как и другие представители своего семейства, тираннозавр был двуногим хищником с массивным черепом, который уравновешивался длинным тяжёлым и жёстким хвостом. По сравнению с большими и мощными задними конечностями этого ящера его передние лапы были совсем небольшими, но необыкновенно могучими для своего размера, и имели два когтистых пальца. Является крупнейшим видом своего семейства, одним из самых больших представителей тероподов и одним из самых крупных наземных хищников за всю историю Земли. Крупнейший из известных полных скелетов этого динозавра имеет длину 12,3 метра[1], высоту до бедра — 4 метра[1], а масса этой особи по оценкам экспертов при жизни достигала 6,8 тонны[1]. Как самое большое плотоядное в своей экосистеме, тираннозавр, скорее всего, был сверххищником и охотился на гадрозавров, цератопсов и, возможно, даже на зауроподов[1], хотя некоторые исследователи предполагают, что питался он преимущественно падалью. Более того, спор о том, являлся ли тираннозавр полноценным охотником и сверххищником либо падальщиком, был одним из самых напряжённых и затяжных в истории палеонтологии. Сейчас большинство специалистов считает, что тираннозавр был хищником-оппортунистом — мог как охотиться, так и питаться падалью[1].
В общей сложности было идентифицировано более 30 скелетов тираннозавров, некоторые из них сохранились почти полностью. Изобилие материала позволило провести углублённые исследования многих аспектов биологии этого животного, включая жизненную историю и даже биомеханику. Особенности питания, физиология и скорость движения — это только немногие из предметов научных споров. Таксономия этого рода также является предметом дискуссий — к примеру, некоторые специалисты относят к нему только один вид (T. rex), а некоторые включают в этот род и тарбозавра, известного из меловых отложений Монголии. Иногда с тираннозавром синонимизируют и некоторые роды североамериканских тираннозавридов. Тираннозавр обрёл большую известность в популярной культуре в качестве свирепого хищника после успеха фильма «Парк юрского периода».
Описание
Размеры
Tyrannosaurus rex был одним из самых крупных наземных хищников всех времён. Самый большой из полных скелетов, FMNH PR2081 по прозвищу «Сью», в длину достигает 12,3 метра[1], а в высоту — 4 метра[1]. Оценка массы в течение многих лет сильно изменялась: от более чем 7,2 тонны[1] до менее чем 4,5 тонны[1][1], самая современная же оценка колеблется между 5,4 тонны и 6,8 тонны[1][1][1][1]. В 2009 году палеонтолог Пакард и его коллеги протестировали метод измерения массы динозавров на слонах и пришёл к выводу, что прежние техники измерения были весьма несовершенны и масса динозавров зачастую переоценивалась, соответственно, масса тираннозавра могла оказаться гораздо меньше, чем было принято считать[1]. Другие же измерения (выполненные при помощи метода Шаблон:Comment, в котором объём животного вычисляется с помощью построения множества сечений и умножения их средней площади на длину тела) позволили заключить, что масса тела самого большого из найденных типовых экземпляров приближается[1] либо даже превышает 9 тонн[1].
Телосложение
Шея тираннозавра, как и у других теропод, имела S-образную форму, но была короткой и мускулистой, что позволяло ей удерживать массивную голову. Передние конечности имели только два когтистых пальца[1] вместе с маленькой пястной костью, рудиментом третьего пальца[1]. Задние конечности, напротив, были самыми длинными (по отношению к телу) среди всех теропод. Позвоночник образован 10 шейными, 12 грудными, пятью крестцовыми и около 40 хвостовыми позвонками. Хвост был тяжёлым и длинным, поскольку выполнял функцию балансира, уравновешивая массивную голову и грузное туловище. Чтобы компенсировать невероятную громоздкость животного, многие кости скелета были полыми, тем самым снижая свой вес, и при этом лишь незначительно уменьшая прочность[1].
Череп
Самый большой из когда-либо найденных черепов тираннозавра в длину достигает полутора метров[1]. Большие окна в черепе способствовали уменьшению его веса и создавали необходимое пространство для прикрепления мышц, как и у всех плотоядных динозавров. Но в других отношениях череп тираннозавра очень отличался от черепов больших теропод, не относящихся к тираннозавридам. Его задняя часть была очень широкой, но рыло было узким, благодаря чему ящер обладал высокоразвитым бинокулярным зрением[1][1]. Черепные кости были тяжёлыми. Носовая и некоторые другие кости были объединены, что предотвращало попадание чего-либо постороннего в пространство между ними. Они были пневматизированы (воздухоносны, то есть в черепе имелись околоносовые пазухи, как и у других нептичьих динозавров). Вероятно, это делало кости более гибкими и лёгкими. Эти и некоторые другие свойства, способствовавшие укреплению черепа, принадлежат к тенденции тираннозавридов к увеличению силы укуса, которая у этих ящеров с огромным преимуществом превосходила таковую у всех теропод-нетираннозаврид[1][1][1]. Конец верхней челюсти имел U-образную форму (в то время как у большинства хищников, не относящихся к тираннозавридам, он был V-образным); такая форма позволяла увеличить количество тканей и костей, которые тираннозавр мог за один укус вырвать из тела жертвы, а также усиливала давление, оказываемое передними зубами ящера[1][1].
Более того, исследования 2012 года, проведённые палеонтологами Карлом Бейтсом и Питером Фолкингемом, позволили предположить, что сила укуса тираннозавра была самой большой среди таковой у всех когда-либо живших наземных животных. Судя по расчётам, задние зубы взрослого T. rex могли смыкаться с силой от 35 до 37 килоньютонов, что в 15 раз больше, чем у африканского льва, в три с половиной раза больше, чем у современного рекордсмена — австралийского гребнистого крокодила и почти в семь раз больше, чем предполагаемая сила укуса аллозавра[1][1]. Ранее, в 2003 году, Мэйсон Б. Меерс из Шаблон:Не переведено 5 получил ещё более высокие результаты. В своих расчётах Меерс оценил силу укуса тираннозавра примерно в 183—235 килоньютонов, что даже превышает оценки силы укуса для самых больших особей мегалодонов[1][1].
У тираннозавра хорошо выражен гетеродонтизм — наличие нескольких типов зубов, различавшихся по форме и функциям[1][1]. Зубы, располагающиеся на передней стороне верхней челюсти, плотно прилегают друг к другу, имеют D-образное поперечное сечение, долотообразное лезвие, укрепляющие гребни и изгиб внутрь. Эти особенности понижали риск слома зуба во время того, как тираннозавр кусал и тащил жертву. Другие зубы более крепкие и массивные, по форме они больше напоминают бананы, чем кинжалы, они шире расставлены и тоже имеют укрепляющие гребни[1]. Зубы верхней челюсти больше зубов нижней (кроме тех, что находятся в её задней части). Самый большой из пока что найденных зубов вместе с корнем достигал высоты 30 сантиметров. Это крупнейший из когда-либо найденных зубов плотоядных ящеров[1].
Классификация
Тираннозавр — типовой род надсемейства тираннозавроидов, семейства тираннозавридов и подсемейства тираннозаврин. Среди других представителей семейства тираннозавридов можно выделить дасплетозавра из Северной Америки и тарбозавра из Монголии[1][1], оба этих рода иногда считают синонимами тираннозавра[1]. Раньше было принято считать тираннозавридов потомками более ранних крупных плотоядных, таких, как Spinosauroidea (мегалозавриды) и карнозавры, но недавно они были признаны потомками более мелких целурозавров[1].
В 1955 году советский палеонтолог Евгений Малеев открыл в позднемеловых отложениях Монголии новый вид, который он назвал Tyrannosaurus bataar[1]. К 1965-му он был переименован в Tarbosaurus bataar[1]. Несмотря на переименование, многие филогенетические исследования показывали, что это был сестринский по отношению к тираннозавру таксон[1], и его часто относили к этому роду[1][1][1]. Недавнее переописание черепа Tarbosaurus bataar показало, что он был гораздо уже, чем у T. rex, а при укусе распределение механического напряжения в черепе было совсем иным — скорее, как у алиорама, другого азиатского теропода[1]. Недавнее кладистическое исследование выявило, что, возможно, не тираннозавр, а именно алиорам, является сестринским таксоном Tarbosaurus bataar, и что если эта гипотеза верна, то Tarbosaurus bataar и тираннозавр в таковых отношениях не находятся и должны рассматриваться в разных родах[1].
Другие окаменелости тираннозавров, найденные в тех же формациях, что и T.rex, были изначально классифицированы как отдельные таксоны, в числе них Aublysodon и Albertosaurus megagracilis,[1] последнему в 1995-м дали название Dinotyrannus megagracilis[1]. Но сейчас эти окаменелости принято считать принадлежащими ювенильным (незрелым) особям T.rex[1]. Маленький, но почти полный череп из Монтаны, длиной в 60 сантиметров — быть может, исключение. Изначально он был приписан одному из видов горгозавра, G. lancensis, Чарльзом Уитни Гилмором в 1946-м году[1], но позднее был отнесен к новому виду — нанотираннусу[1]. Мнения учёных разделяются насчет того, имеет ли таксон N. lancensis место быть. Многие ученые считают, что это тоже череп незрелого тираннозавра[1]. Различия этих двух видов невелики (в частности, у N. lancensis больше зубов). Поэтому некоторые специалисты рекомендуют оставить их как отдельные таксоны, пока дальнейшие исследования не прольют свет на этот вопрос[1][1].
Палеобиология
История жизни
Исследование нескольких экземпляров ювенильных особей тираннозавров позволило описать онтогенетические изменения, оценить продолжительность жизни и скорость роста. Самая маленькая из когда-либо найденных особей, (LACM 28471, известная как «Шаблон:Comment теропод») имела массу тела лишь 30 килограммов, в то время как самая большая, (FMNH PR2081, по прозвищу «Сью») весила более 5400 килограммов. Гистологический анализ костей тираннозавра показал, что на момент смерти «Джорданский теропод» был двухлетнего возраста, в то время как «Сью» было 28 лет, этот возраст, вероятно, был близок к максимально возможному для данного вида[1].
Файл:Tyrantgraph.png График, показывающий предполагаемую кривую роста тираннозавра, на оси
абсцисс — возраст особей, на оси
ординат — масса тела (кривая роста для тираннозавра обозначена чёрным цветом, для сравнения приведены кривые роста у других тираннозаврид). Основано на исследованиях Эриксона и других
2004 года Гистологические исследования позволили определить возраст и других экземпляров. Чтобы показать зависимость массы той или иной найденной особи тираннозавра от её возраста, можно изобразить график — «кривую роста» (Шаблон:Lang-en). У T. rex кривая роста имеет S-образную форму, причём быстрое увеличение массы начинается около 14 лет (этому возрасту соответствует масса 1800 кг). В течение этой стремительной фазы роста тираннозавр на протяжении 4 лет набирает по 600 килограммов в год. Когда особь достигает восемнадцатилетнего возраста, её рост быстро замедляется. К примеру: у 28-летней особи по прозвищу «Сью» и 22-летней канадской особи (RTMP 81.12.1) разница в весе составляла лишь 600 килограммов[1]. Другие недавние гистологические исследования, проведённые различными учёными, подтверждают эти результаты, показывая, что стремительный рост начинает замедляться при достижении шестнадцатилетнего возраста[1]. А эти исследования, в свою очередь, подкреплены некоторыми другими, которые, тем не менее, показывают, что рост идёт гораздо быстрее, и за счет него животное набирает 1800 килограммов. Хотя все эти оценки значительно превышали результаты, полученные ранее, их авторы заметили, что данные результаты существенно сгладили различие между настоящими показателями роста и теми, что изначально ожидали от животного таких размеров[1]. Внезапное изменение скорости роста в конце этой «вспышки», вероятно, говорит о достижении физической зрелости — в пользу этой гипотезы говорит обнаружение медуллярной ткани в бедренной кости особи возрастом от 16 до 20 лет из Монтаны (MOR 1125, также известна как «Би-рекс», (Шаблон:Lang-en)). Медуллярная ткань — это костная ткань, откладывающаяся у самок птиц в полостях костей в период овуляции и служащая резервуаром кальция[1]. Наличие её у «Би-рекс» позволяет утверждать, что она была особью репродуктивного возраста[1]. Дальнейшие исследования показали, что этой особи было 18 лет[1]. У других тираннозавридов кривые роста имеют чрезвычайно похожий вид, несмотря на то, что в связи с меньшими размерами взрослых особей скорость роста у них ниже[1].
Более половины найденных скелетов T. rex принадлежали особям, умершим в течение шести лет после достижения половозрелого возраста, что также характерно для других тираннозавридов и некоторых больших, долгоживущих млекопитающих и птиц в наши дни. Эти виды характеризуются высокой смертностью детёнышей, но среди ювенильных особей, однако, смертность относительно низка. Среди половозрелых особей уровень смертности опять возрастает, что связано со стрессом при размножении. В ходе одного исследования было высказано предположение, что редкость нахождения ювенильных особей связана с их низкой смертностью. Кроме того, эта редкость может быть связана с неполнотой окаменелого материала или же с пристрастием коллекционеров окаменелостей к более крупным, зрелищным экземплярам[1]. В своей лекции 2013 года палеонтолог Шаблон:Нп5 предположил, что динозавры «жили быстро и умирали молодыми», так как быстро размножались, в отличие от млекопитающих, у которых продолжительность жизни больше в связи с тем, что они больше времени затрачивают на продолжение рода[1]. Палеонтолог Грегори С. Пол также пишет, что тираннозавры быстро размножались и умирали в молодом возрасте, но их малую продолжительность жизни объясняет тем, что жили они слишком опасной жизнью[1].
Половой диморфизм
Когда число найденных экземпляров стало увеличиваться, учёные начали анализировать различия между особями и установили два определённых типа телосложения («морфы»), что также было свойственно другим видам теропод. Один из этих типов телосложения относился к более крепким и массивным особям (таких особей стали именовать «робустными»), в то время как животных с другим типом телосложения — «грацильными», «стройными». Некоторые морфологические различия между двумя этими типами были применены для того, чтобы исследовать половой диморфизм у тираннозавров, причём «робустных» особей обычно считали самками. К примеру, таз нескольких робустных экземпляров расширен, что было связано, возможно, с необходимостью откладывать яйца[1]. Также предполагалось, что одной из главных особенностей морфологии «робустных» особей была редукция шеврона первого хвостового позвонка, связанная с необходимостью выведения яиц из репродуктивного тракта (эта особенность была ошибочно приписана крокодилам)[1].
В последние годы доказательства полового диморфизма стали сходить на нет. Исследования 2005 года показали, что прежние данные о половом диморфизме в строении шевронов позвонков крокодилов были ошибочными, что ставит под сомнение наличие подобной особенности у тираннозавров[1]. Шеврон полных размеров был обнаружен на первом хвостовом позвонке «Сью» — чрезвычайно крепкой особи, что говорит о том, что данный признак характерен не только для одного типа телосложения. Поскольку скелеты T. rex были обнаружены на просторах от Саскачевана до Нью-Мексико, различия в анатомии особей могут быть вызваны скорее географической изменчивостью, чем половым диморфизмом. Также они могут быть связаны с разным возрастом животных — «робустные» скелеты могли принадлежать старым особям[1].
Точно определить пол удалось только у одного экземпляра T. rex. Изучение останков «Би-рекса» позволило выявить наличие мягких тканей в нескольких костях. Некоторые из этих тканей были идентифицированы как медуллярные, то есть специальные ткани, присутствующие у современных птиц в течение овуляции в качестве источника кальция, необходимого для образования яичной скорлупы. Поскольку яйца откладывают только самки птиц, медуллярную ткань обычно находят в костях особей женского пола, хотя она может образовываться и у самцов при введении таких женских репродуктивных гормонов, как эстрогены. На основании всего этого твердо считается, что «Би-рекс» является особью женского пола, погибшей в период овуляции[1]. Недавние исследования показали, что медуллярной ткани никогда не бывает у крокодилов, которые считаются ближайшими современными родственниками динозавров наряду с птицами. Общность наличия медуллярной ткани у птиц и тероподов является ещё одним свидетельством их эволюционной близости[1].
Осанка
Современные изображения в музеях, живописи и фильмах демонстрируют T. rex с телом, примерно параллельным земной поверхности и хвостом, вытянутым позади тела и уравновешивающим тяжёлую голову[1].
Как и многих двуногих динозавров, тираннозавров издавна изображали как «живой штатив», с телом, на 45° или меньше отклоняющимся от вертикальной позиции и хвостом, волочащимся вдоль земли, примерно как у кенгуру. Этому мнению положил начало Джозеф Лейди, выполнивший реконструкцию гадрозавра в 1865 году — первый рисунок, запечатлевший динозавра как двуногого существа[1]. В 1915 году, когда уже было принято считать тираннозавра двуногим, известный учёный Генри Файрфилд Осборн (бывший президент AMNH — Американского музея естественной истории, Нью-Йорк), ещё раз подтвердил эту точку зрения, открыв первый полный скелет тираннозавра, который запечатлел животного именно в такой позе. В двуногой позе этот скелет простоял целых 77 лет, вплоть до того момента, как в 1992 году был перемонтирован[1].
К 1970 году учёные поняли, что предположение о такой позе неверно и что она не могла сохраняться ящером при жизни, иначе это привело бы к вывиху или ослаблению нескольких суставов, включая бедренный и сочленение между головой и позвоночным столбом[1]. Неправильно установленный в AMNH скелет послужил основой для подобных изображений ящера во многих фильмах и произведениях живописи (таких, как знаменитая фреска «Век рептилий» Рудольфа Франца Цаллингера, которую можно увидеть в Шаблон:Нп5 Йельского университета)[1], созданных до 1990-х, когда в таких фильмах, как «Парк юрского периода», была представлена уже более правильная осанка ящера[1].
Передние конечности
Когда T. rex был обнаружен, единственной известной частью его передних конечностей была плечевая кость[1]. Во впервые установленном в музее скелете, который был показан публике в 1915 году, вместо ещё не обнаруженных конечностей тираннозавра Осборн поставил более длинные, трёхпалые лапы, похожие на таковые у аллозавра[1]. Годом ранее Лоуренс Ламб описал короткие, двупалые передние лапы близкого родственника T. rex горгозавра[1]. Это давало сильные основания утверждать, что у T. rex передние конечности были похожими, однако это предположение было доказано только в 1989 году, когда были идентифицированы полные окаменелые останки передних лап тираннозавра (экземпляр MOR 555, известный как «Уонкел рекс» — Шаблон:Lang-en)[1]. В скелете «Сью», кстати говоря, передние конечности тоже полностью сохранились[1]. Передние конечности тираннозавра чрезвычайно малы по отношению к общим размерам тела, в длину они достигают лишь одного метра, некоторые учёные вообще считают их рудиментарными органами. Но на их костях есть большие участки для прикрепления мышц, что говорит о значительной силе. Впервые эта особенность была замечена ещё в 1906 году Осборном, который предположил, что передние конечности могли использоваться для удержания полового партнёра во время спаривания[1]. Также предполагалось, что они использовались с целью помочь животному подняться из лежачего положения[1]. Другой их функцией могло быть удержание пытающейся вырваться жертвы, пока хищник убивал её своими огромными челюстями. В пользу этого говорят некоторые биомеханические признаки.
Кости передних конечностей T. rex отличаются исключительно толстым поверхностным (непористым) слоем, что говорит об их способности выдерживать большую нагрузку. Двуглавая мышца плеча взрослого тираннозавра была способна самостоятельно поднять груз весом в 200 килограммов[1][1]. Другие мышцы, такие, как плечевая, работали параллельно с двуглавой, чтобы сгибание локтя становилось ещё мощнее. Бицепс T. rex был в три с половиной раза мощнее человеческого. Диапазон движений предплечья тираннозавра был ограничен, плечевые и локтевые суставы могли двигаться лишь на 40 и 45 градусов соответственно. Для сравнения: те же два сустава дейнониха могли передвигаться на 88 и 130 градусов, а у человека — на 360 и 165 градусов соответственно. Массивность костей передних лап, мускульная сила и ограниченный диапазон движений — всё это говорит о наличии у ящера особой системы, развившейся для того, чтобы крепко удерживать жертву, несмотря на все её отчаянные усилия при попытке вырваться[1].
Мягкие ткани
В марте 2005 года в выпуске журнала Science Мэри Хигби Швейцер, палеонтолог из Университета штата Северная Каролина, и её коллеги объявили о находке мягких тканей в костном мозге окаменелой ноги T. rex. Эта кость была намеренно разбита для вертолётной перевозки и поэтому подошла для обследования на наличие мягких тканей[1]. Обозначенный как MOR 1125 (1125-й экземпляр в коллекции Музея Скалистых гор), этот динозавр был найден в формации Хелл-Крик. Специалисты смогли различить гибкие разветвлённые кровеносные сосуды и волокнистую, но эластичную костную матрицу. Более того, внутри матрицы и сосудов были найдены микроструктуры, напоминающие кровяные тельца. Все эти объекты очень похожи на кровяные тельца и сосуды страусов. Исследователям пока что неизвестно, сохранился ли данный материал вследствие какого-то процесса, отличного от обычной фоссилизации, либо же это ткани в исходном виде, и пока они воздерживаются от каких-либо предположений на этот счёт[1]. Если это ткани в исходном виде, то любые сохранившиеся белки могут позволить расшифровать часть структуры ДНК динозавров, поскольку каждый белок, как известно, закодирован собственным геном. Отсутствие предыдущих подобных находок, вероятно, объясняется тем, что специалисты ранее просто не верили в то, что до наших дней могут дойти живые ткани, и, соответственно, не занимались поиском и исследованием таковых. Со времени первого подобного открытия учёные обнаружили ещё два экземпляра тираннозавров и один экземпляр гадрозавра, у которых также сохранились тканевые структуры такого рода[1]. Исследования некоторых из этих тканей позволили предположить, что птицы родственны тираннозаврам более, чем каким-либо современным животным[1].
В ходе исследований, о которых сообщалось в журнале Science в апреле 2007 года, Асара с коллегами заключили, что коллаген, обнаруженный в очищенной кости T. rex, ближе всего к коллагену курицы и более далёк от коллагена лягушек и тритонов. Открытие белков в окаменелостях возрастом в десятки миллионов лет, как и в костях мастодонта возрастом 160 тыс. лет, может перевернуть все традиционные взгляды на останки древних животных и заставить палеонтологов переключиться с охоты за окаменелостями на биохимию. До этих открытий было принято считать, что в течение процесса фоссилизации все живые ткани заменялись инертными минералами. Палеонтолог Ганс Ларссон из Мак-Джильского университета города Монреаль, не принимавший участия в этих исследованиях, назвал данные открытия «вехами» и предположил, что динозавры могут «войти в сферу молекулярной биологии и запустить палеонтологию в современный мир»[1].
Предполагаемое наличие мягких тканей у тираннозавра было подвергнуто сомнению палеонтологом Томасом Кеем (Шаблон:Lang-en) из Вашингтонского университета и его коллегами в 2008 году. Они заявили, что в действительности внутри кости тираннозавра находилась слизистая биоплёнка, созданная бактериями, покрывавшими стенки полостей, где раньше были кровеносные сосуды и кровяные тельца[1]. Исследователи также посчитали то, что ранее ввиду содержания железа было идентифицировано как остатки кровяных телец, фрамбоидами, микроскопическими минеральными сферами, содержащими железо. Подобные сферы находили и у множества других окаменелостей из разных периодов, в том числе у аммонитов. Что касается последних, то у них фрамбоиды нашли в том месте, где содержащееся в них железо не могло иметь никакого отношения к наличию крови[1]. Швейцер подвергла доводы Кея жёсткой критике, заявляя, что нет должных свидетельств тому, что биоплёнка способна образовывать разветвления и полые трубки, которые она обнаружила в ходе своих исследований. В 2011 году Сан Антонио и Швейцер вместе со своими коллегами опубликовали анализ тех частей коллагена, что были обнаружены, считая сохранившиеся образования внутренними частями коллагеновых колец постольку, поскольку именно их формирование происходит в ходе долгого периода распада белков[1]. Другое исследование ставит под сомнение идентификацию тех мягких тканей как биоплёнок и подтверждает наличие внутри кости «ветвящихся, похожих на сосуды структур»[1].
Кожа и перья
Шаблон:Main
Многие учёные считают, что T. rex имел перья (по крайней мере, на отдельных частях тела), хотя прямых подтверждений этого нет[1]. Это предположение основывается на наличии перьев у родственных видов схожего размера. Доктор Марк Норелл из Американского музея естественной истории подытожил все факты и свидетельства следующими словами: «У нас столько же свидетельств того, что T. rex был покрыт перьями, хотя бы на определённых этапах своей жизни, сколько и того, что австралопитеки, такие как Люси, были покрыты шерстью»[1].
Первое свидетельство наличия перьев у тираннозавроидов было обнаружено у маленького динозавра Dilong paradoxus, чьи останки были найдены в знаменитой китайской формации Исянь, о чём было сообщено в журнале Nature в 2004 году. Как и у многих других тероподов из той формации, окаменелый скелет этого существа был окаймлён слоем нитевидных структур, которые обычно считают протооперением[1]. Поскольку у более крупных тираннозавроидов нашли окаменевшую чешую, учёные, изучавшие динозавра Dilong, сделали вывод, что количество перьев с возрастом уменьшалось — незрелые особи были оперены, а к зрелому возрасту, когда животные достигали больших размеров, у них оставалась лишь чешуя, так как перья больше не требовались для сохранения тепла[1]. Тем не менее, последующие открытия подвергли эту гипотезу сомнению: даже у больших тираннозавроидов нашли перья, покрывавшие большую часть тела[1].
В то время как «Wyrex» (экземпляр BHI 6230) — кусок окаменевшей кожи, найденный в Монтане в 2002 году, а также некоторые другие экземпляры гигантских тираннозавроид показывают наличие чешуи[1], у других (таких, как Yutyrannus huali, который достигал 9 метров в длину и весил 1,4 тонны) сохранились отпечатки перьев на разных участках тела. В связи с этим можно утверждать, что перья могли целиком покрывать их тела[1]. Возможно, что количество перьев и характер перьевого покрова у тираннозавроид могли изменяться со временем, в связи с изменением размеров ящеров, потеплением климата или другими факторами[1].
Терморегуляция
Тираннозавра, как и большинство других динозавров, долгое время считали холоднокровным. Гипотеза о холоднокровности динозавров была подвергнута сомнению палеонтологами Робертом Бэккером и Джоном Остромом в конце 1960-х годов[1][1]. Они утверждали, что тираннозавр был теплокровным и вел очень активный образ жизни[1]. В пользу этой гипотезы может говорить высокий темп роста тираннозавров, сопоставимый с таковым у млекопитающих и птиц. Графики (кривые роста) показывают, что рост этих ящеров приостанавливался ещё в неполовозрелом возрасте, в отличие от большинства других позвоночных[1].
Соотношение изотопов кислорода в костях тираннозавров позволяет определить температуру, при которой эти кости росли. У одного экземпляра температура позвоночника и берцовой кости отличалась не более чем на 4—5 °C. По мнению палеонтолога Риза Баррика и геохимика Уильяма Шоуэрса, маленькая разница в температуре туловища и конечностей говорит о том, что тираннозавр поддерживал постоянную внутреннюю температуру тела (Шаблон:Нп3), и что его метаболизм — нечто среднее между метаболизмами холоднокровных рептилий и теплокровных млекопитающих[1]. Однако другие учёные считают, что нынешнее соотношение изотопов кислорода в окаменелостях не соответствует изначальному, поскольку оно могло изменяться во время или после фоссилизации (это явление называют диагенезом)[1]. Тем не менее, Баррик и Шоуэрс нашли аргумент в пользу своих утверждений: в останках гиганотозавра — теропода, жившего на другом континенте и на десятки миллионов лет ранее тираннозавра, были обнаружены такие же показатели[1]. У птицетазовых динозавров также находили свидетельства гомеотермии, в то время как у ящериц из семейства варановых из той же формации ничего подобного обнаружено не было[1]. Но даже если тираннозавр действительно поддерживал постоянную температуру тела, это не означает, что он точно был теплокровным. Такая терморегуляция может быть объяснена Шаблон:Нп3, которая наблюдается у ныне живущих кожистых морских черепах[1][1].
Следы
Тираннозавру рекс были приписаны два отдельных отпечатка стопы. Один из них обнаружил геолог Чарльз Пиллмор в 1983 году на Шаблон:Нп3, в штате Нью-Мексико. Первоначально след был приписан гадрозавриду, однако большой размер пятки и четвёртый палец, не характерные для орнитопод, натолкнули на вывод о принадлежности отпечатка тираннозавру. В 1994 году Мартин Локли и Адриан Хант дали этому ихнотаксону название Tyrannosauripus pillmorei. След был оставлен в водно-болотных угодьях. Длина отпечатка равняется 83 сантиметрам, а ширина — 71 сантиметру[1].
О втором вероятном отпечатке стопы тираннозавра было впервые упомянуто в 2007 году британским палеонтологом Филом Мэннингом, найден он был в формации Хелл-Крик, штат Монтана. Длина этого следа составляет 72 сантиметра. Точно не известно, принадлежал ли след тираннозавру, поскольку в отложениях Хелл-Крик нашли останки двух крупных тероподов — собственно тираннозавра и нанотираннуса. Дальнейшие исследования следа из Хелл-Крик будут направлены на его сравнение с отпечатком из Нью-Мексико.
Передвижение
Два основных вопроса, касающихся передвижения тираннозавра: насколько быстро он мог разворачиваться и какова была его максимальная скорость при прямолинейном движении. Оба связаны с вопросом о том, был ли этот ящер охотником или падальщиком.
Скорее всего, тираннозавр разворачивался медленно, вероятно, для поворота на 45° ему требовалась одна или две секунды[1]. Причина этих сложностей заключается в том, что большая часть массы тираннозавра была удалена от его центра тяжести, однако он мог уменьшить это расстояние, выгибая спину и хвост и прижимая голову и конечности к туловищу[1].
Существует много предположительных оценок максимальной скорости тираннозавра, в основном около 39,6 км/ч (11 м/с), наименьшая оценка — от 18 км/ч (5 м/с), а наивысшая — 72 км/ч (20 м/с). Исследователи должны полагаться на различные технологии, поскольку было найдено множество дорожек следов, оставленных большими тероподами во время хождения, но не было найдено ни одной, оставленной ими во время бега. Это может означать, что к бегу они не были способны[1]. Ученые, считающие, что тираннозавр мог бегать, объясняют это тем, что полые кости и некоторые другие свойства значительно облегчали тело. Кроме того, некоторые другие специалисты отмечали, что мышцы ног тираннозавра были больше, чем у любого современного животного, благодаря чему он мог развивать большую скорость в 40—70 километров в час[1].
В 1993 году Джек Хорнер и Дон Лессем заявили, что тираннозавр был медлительным животным и, вероятно, не умел бегать, поскольку отношение длины бедренной кости к длины берцовой превышало 1, как и у большинства крупных тероподов и современного слона[1]. Тем не менее, в 1998 году Томас Хольтц отмечал, что у тираннозавридов и родственных им групп дистальные (дальние) компоненты задних конечностей были значительно длиннее по отношению к бедренной кости, чем у большинства других теропод, а также что у тираннозавридов и их родственников плюсна очень плотно сцеплялась с другими частями стопы, благодаря чему более эффективно передавала усилие (импульс) от стопы к голени, чем то было у более ранних теропод. Хольтц заключил, что вследствие этого тираннозавриды и родственные им динозавры были самыми быстрыми из всех больших теропод[1]. Эти предположения он упомянул и в своей лекции 2013 года, утверждая, что у аллозавров при том же размере туловища, что и у тираннозавров, ступни были короче[1].
В 1998 году Кристиансен предположил, что кости ног тираннозавра были не сильнее, чем кости ног слона, скорость которых весьма ограниченна и которые не позволяют бегать. Из этого он сделал вывод, что максимальная скорость тираннозавра не превышала 39,6 км/ч (11 м/с, это примерно равняется скорости человека-спринтера). Однако он также заметил, что справедливость этого вывода зависит от правильности многих поставленных под сомнение гипотез[1].
В 1995 Фарлоу со своими коллегами оценил массу тираннозавра от 5,4 до 7,3 тонны. Для такого тяжелого животного падение во время быстрого движения могло оказаться роковым[1]. Тем не менее, жирафы способны развивать скорость до 50 км/ч, рискуя сломать ногу или получить ещё худшие повреждения, которые могут привести к смерти не только в дикой среде, но и в безопасной среде — например, зоопарке[1][1]. Весьма вероятно, что в случае необходимости тираннозавр тоже подвергал себя подобному риску[1][1].
В своем исследовании, опубликованном в журнале «Гайа» (Шаблон:Lang-en), Грегори С. Пол заявил, что тираннозавр был гораздо лучше приспособлен к бегу, чем люди или слоны, поскольку имел большую подвздошную кость и большой гребень большеберцовой кости, на которых, соответственно, крепились большие мышцы. Также Пол отмечал, что формула для расчета скорости по силе костей, выведенная палеонтологом Александром в 1989 году, была лишь частично верной. По его мнению, в формуле чрезмерно большое значение уделялось длине костей, а кости значительной длины согласно этой формуле получались неестественно слабыми. Также он утверждал, что риск получить раны в схватках с другими животными был вполне сопоставим с риском упасть во время бега[1].
Недавние исследования особенностей движения тираннозавра показывают, что бегал этот ящер со средней скоростью, не быстрее чем 40 км/ч. К примеру, в публикации журнала Nature от 2002 года рассказывается о математической модели (применимой также к современным животным: аллигаторам, курицам, эму, страусам, а также человеку[1]), с помощью которой можно оценить размер мышц, необходимых для быстрого бега (свыше 40 км/ч)[1]. Исследователи посчитали, что тираннозавр не мог развивать скорость выше 40 км/ч, поскольку для этого потребовались бы мускулы, составляющие 40—86 % от общей массы тела. Даже умеренно большая скорость потребовала бы больших мышц. Однако этот вопрос разрешить трудно, поскольку неизвестно, насколько большие мускулы имел тираннозавр. Если они были небольшими, тираннозавр мог лишь ходить или легко бегать со скоростью 18 км/ч[1].
В исследовании 2007 года использовалась компьютерная модель для измерения скорости бега, основывалась она на информации, полученной непосредственно из окаменелостей. Согласно ей, максимальная скорость тираннозавра достигала 29 км/ч (8 м/с). Скорость среднего футболиста чуть меньше, а спринтер может развивать скорость 43 км/ч (12 м/с). Примечательно, что максимальную скорость трехкилограммовой (возможно, ювенильной[1]) особи компсогната модель оценила в 64 км/ч (17,8 м/с)[1][1].
Тем не менее, в 2010 году выпускник университета Альберты Скотт Персонс предположил, что скорость тираннозавра могла увеличиваться с помощью сильных мышц хвоста[1]. Он выяснил, что у теропод, таких, как тираннозавр, расположение некоторых мускулов отличалось от такового у современных млекопитающих и птиц, но имело некоторые сходства с мышцами современных рептилий[1]. Студент сделал вывод, что хвостово-бедренные мышцы (Шаблон:Lang-lat) повышали способность к бегу и проворство тираннозавра, помогали сохранять равновесие, а также были наиболее важными сгибающими мышцами ног[1]. Исследование также показало, что в скелете тираннозавра и родственных ему тероподов были такие приспособления, как Шаблон:Нп3, которые способствовали увеличению мускулов хвоста. Также было выявлено, что масса хвостовых мышц тираннозавра составляла 25—45 % от всей мышечной массы, а масса хвостово-бедренной мышцы составляла 58 % от массы всех хвостовых мышц. Согласно Персонсу, увеличение мышц хвоста смещало центр массы ближе к задним конечностям, что помогало им лучше выдерживать тяжесть тела и сохранять равновесие. Помимо этого Персонс отмечает, что хвост тираннозавра был богат сухожилиями и септами (перегородками), что придавало ему эластичность и, следовательно, улучшало движение. Исследователь добавляет, что у нептичьих тероподов хвосты были шире, чем изображали ранее, и были столь же или даже более широкими в боковом направлении, чем в спинно-брюшном[1][1].
Генрих Маллисон из Берлинского музея естественной истории в 2011 году представил теорию, согласно которой тираннозавр и многие другие динозавры могли развивать относительно высокую скорость с помощью коротких быстрых шагов (современные млекопитающие и птицы, напротив, во время бега делают большие шаги). Согласно Маллисону, скорость этих динозавров не зависела от силы суставов и не требовала дополнительной мышечной массы на ногах (в частности, на лодыжках). В подтверждение своей теории Маллисон изучил конечности разных динозавров и установил, что они отличались от конечностей современных млекопитающих и птиц, поскольку особое строение скелета строго ограничивало длину их шага, но зато они имели относительно большие мускулы на крупе. Тем не менее, он нашел несколько сходств между мускулатурами динозавров и быстрых современных животных: на их крупе мышечная масса больше, чем на лодыжках. Маллисон также предполагает, что отличия между динозаврами и ныне живущими млекопитающими и птицами позволят составить уравнения для расчета скорости по длине шага. Однако Джон Хатчисон считает, что правильнее сначала изучить мускулы динозавров и выявить, как часто они могли сокращаться[1][1].
Учёные, считающие, что тираннозавр был неспособен бегать, оценивают максимальную скорость ящера примерно в 17 км/ч. В любом случае, это превышает скорость добычи тираннозавра — анкилозаврид и цератопсов[1]. Более того, сторонники мнения, что тираннозавр был активным хищником, утверждают, что скорость бега тираннозавра несущественна — даже если он был медлительным, все равно передвигался быстрее своих жертв[1]. Томас Хольтц отмечал, что ступня тираннозавра была пропорционально длиннее, чем у тех животных, на которых он охотился — утконосых и рогатых динозавров[1]. Тем не менее, в 2000 году Пол и Кристиансен возразили, что поздние цератопсы имели прямые передние конечности, и их крупные виды могли передвигаться так же быстро, как современные носороги[1]. Однако против их довода говорят отметины от зубов тираннозавра на окаменелостях цератопсов, причем некоторые из них были нанесены во время атаки на живых цератопсов (см. ниже), то есть тираннозавр был достаточно быстр, чтобы поймать свою добычу[1].
Мозг и органы чувств
Исследования, проведенные под руководством Лоренса Уитмера и Райана Риджли, палеонтологов из университета Огайо, добавили некоторые подробности к уже известным фактам об органах чувств у тираннозавра. Специалисты заявили, что разделяют мнение о повышенных сенсорных способностях целурозавров, подчеркнули быстрые и хорошо скоординированные движения зрачков и головы, а также способность улавливать низкочастотные звуки, которые помогали тираннозавру обнаруживать добычу на больших расстояниях, и превосходное обоняние[1]. Исследование Кена Стивенса из университета Орегона позволило сделать вывод, что у тираннозавра было острое зрение. Стивенс выявил, что бинокулярный диапазон у тираннозавра равнялся 55 градусам — больше чем у современного ястреба. Также было установлена, что острота зрения тираннозавра в 13 раз превышала таковую у человека, и, соответственно, превышала остроту зрения орла (которая лишь в 3,6 раза выше человеческой). Все это позволяло тираннозавру распознавать Шаблон:Прояснить[1][1][1][1].
Томас Хольтц-младший отмечал, что повышенное глубинное восприятие тираннозавра могло быть связано с животными, на которых он охотился; таковыми являлись рогатый динозавр трицератопс, панцирный динозавр анкилозавр и утконосые динозавры, все из которых обладали сложным социальным поведением. В отличие от тираннозавра, акрокантозавр — другой гигантский хищный динозавр — имел весьма ограниченное глубинное восприятие, так как охотился на больших зауропод, которые ко времени появления тираннозавра в большинстве своем исчезли[1].
Тираннозавр примечателен большими размерами Шаблон:Нп3 и обонятельных нервов по отношению к размеру всего мозга, вследствие чего ящер обладал отличным обонянием, и, соответственно, мог учуять запах мертвечины на большом расстоянии. Вероятно, обоняние тираннозавра было сопоставимо с таковым у современных стервятников. Исследование обонятельных луковиц у 21 рода нептичьих динозавров показало, что наиболее развитым обонянием среди них обладал именно тираннозавр[1].
У тираннозавра была очень длинная улитка уха, что нехарактерно для теропод. Длина улитки часто связана с остротой слуха, или, по крайней мере, показывает, насколько важным аспектом поведения был слух. Следовательно, в жизни тираннозавра это чувство играло большую роль. Исследования показали, что тираннозавр наиболее хорошо улавливал низкочастотные звуки[1].
Исследование, проведенное Грантом Р. Херлбертом вместе с вышеупомянутыми Уитмером и Риджли, было направлено на измерение коэффициента энцефализации (EQ) динозавров, основываясь на этом показателе у рептилий (REQ) и птиц (BEQ), также палеонтологи измеряли отношение массы конечного мозга к общей мозговой массе. Ученые установили, что тираннозавр имел самый большой относительный размер мозга среди всех нептичьих динозавров, кроме небольших представителей манирапторов (бэмбираптора, троодона и орнитомима). Однако было выявлено, что относительный размер мозга тираннозавра лежал в пределах диапазона такового у современных рептилий, хотя и существенно выше среднего значения. В частности, отношение массы конечного мозга к общей мозговой массе составляло от 47 до 50 %, что превышает наименьший показатель у птиц, но приближается к типичному показателю у аллигаторов (45,9-47,9 %)[1].
Особенности питания
Спор о том, был ли тираннозавр активным хищником или падальщиком, начался тогда же, когда и спор об особенностях его передвижения. В 1917 году Ламбе описал скелет горгозавра — близкого родственника тираннозавра и заключил, что горгозавр и, соответственно, тираннозавр были исключительно падальщиками, поскольку зубы горгозавра почти не были изношены[1]. Однако в наши дни этот аргумент не воспринимают всерьёз, так как известно, что у тероподов на месте изношенных зубов быстро вырастали новые. Со времени открытия тираннозавра ученым стало ясно, что это был хищник, и как большинство современных крупных плотоядных, он мог питаться падалью или по возможности отнимал добычу у других хищников[1].
Палеонтолог Джек Хорнер отстаивал гипотезу о том, что тираннозавр был исключительно падальщиком и никогда не занимался активной охотой[1][1][1]. Однако он же утверждал, что никогда не публиковал эту гипотезу в научной литературе, а лишь преподносил её широкой публике и использовал в образовательных целях, чтобы научить детей, собирающихся стать учёными, в будущем не допускать ошибок и не делать голословных утверждений (таких, как то, что тираннозавр был активным хищником)[1]. Тем не менее, в научно-популярной литературе Хорнер предоставил следующие доказательства в пользу своего предположения:
- Передние конечности тираннозавра слишком малы по сравнению с таковыми у других хищников. По мнению Хорнера, они были совершенно неспособны к захвату жертвы[1].
- У тираннозавра было превосходное обоняние, как у стервятников, которое позволяло ему обнаруживать туши мёртвых животных на больших расстояниях (см. выше). Оппоненты Хорнера заявили, что сравнение со стервятниками необоснованно, так как единственными чистыми падальщиками среди современных животных являются большие планирующие птицы, способные покрывать большие расстояния и обладающие высокоразвитыми органами чувств[1]. Тем не менее, ученые из Глазго пришли к выводу, что экосистема, столь же плодородная, сколь современная Серенгети, могла обеспечить крупного падальщика большим количеством падали. Однако для подтверждения этого вывода тираннозавр должен был быть холоднокровным, чтобы получить из падали больше калорий, чем он затратил на поиск пропитания. Они также заявили, что на территории современной экосистемы Серенгети нет крупных наземных падальщиков, поскольку они не выдержали бы конкуренции со стороны больших планирующих птиц, в то время как во времена тираннозавров их экологическая ниша пустовала, и у ящеров конкурентов не было[1].
- Зубы тираннозавра могли дробить кости, и поэтому ящер мог получить максимум пропитания из останков мертвых животных. Карен Чин со своими коллегами нашли костные фрагменты в копролитах, которые они приписали тираннозавру, однако заявили, что тираннозавр не обладал способностью пережевывать кости и извлекать костный мозг, как это делают современные гиены[1].
- Поскольку некоторые жертвы тираннозавра могли передвигаться быстро, свидетельства в пользу того, что тираннозавр ходил, а не бегал, говорят о том, что он был падальщиком[1][1]. С другой стороны, недавние исследования показали, что тираннозавр, будучи медлительнее больших наземных хищников современности, мог перемещаться достаточно быстро, чтобы успешно охотиться на крупных гадрозавров и цератопсов[1][1].
Файл:DMNS Edmontosaurus.png Скелет
эдмонтозавра, выставленный в Денверском музее природы и науки. Повреждения на хвосте говорят о том, что эдмонтозавра, вероятно, ранил тираннозавр во время охоты
Другие свидетельства говорят в пользу того, что тираннозавр был активным хищником. Глазницы тираннозавра были расположены так, что взгляд был устремлен вперед, и у ящера было хорошее бинокулярное зрение — даже лучше, чем у ястребов. Хорнер также отмечал, что в родословной тираннозавров прослеживается неуклонное улучшение бинокулярного зрения. Неизвестно, почему имела место эта долгосрочная тенденция, если тираннозавр был исключительным падальщиком (падальщики не нуждаются в повышенном глубинном восприятии)[1][1]. Более того, в современном мире хорошее стереоскопическое зрение присуще именно быстро бегающим хищникам[1].
В Монтане был найден скелет гадрозаврида Edmontosaurus annectens с повреждениями на хвосте, которые были оставлены укусом тираннозавра. По всей видимости, эдмонтозавр выжил после атаки хищника, и повреждения зажили; все это может говорить в пользу того, что тираннозавр охотился на живых динозавров, то есть был активным хищником[1]. Однако, как оценивается, атаковавший эдмонтозавра был молодой особью, с коронкой зуба достигавшей только 3.75 см высоты[1]. Также были найдены окаменевшие свидетельства агрессивных столкновений между тираннозавром и трицератопсом: следы от зубов на надбровном роге и некоторых костях шеи одного экземпляра трицератопса (впоследствии вместо сломанного рога вырос новый, а рана на шее зажила). Однако точно неизвестно, кто в схватке был агрессором[1]. Как сказано выше, окаменелости свидетельствуют о том, что раны трицератопса зажили — из этого можно сделать вывод, что он выжил в битве, вырвался и убежал, или даже победил тираннозавра. Этого мнения придерживается палеонтолог Питер Додсон, считающий, что рога трицератопса могли нанести увечья, смертельные для тираннозавра[1]. Но, в то же время, следы от зубов тираннозавров на костях трицератопсов без следов заживления достаточно распространены[1][1]. И существуют окаменелости, возможно, демонстрирующие удачную охоты куда более мелких тираннозавридов (возможно, молодых тираннозавров) на крупных трицератопсов[1][1].
Во время изучения экземпляра «Сью» палеонтолог Питер Ларсон обнаружил сросшиеся после перелома малоберцовую кость и хвостовые позвонки, а также трещины на лицевых костях и зуб другого тираннозавра, застрявший в шейных позвонках. Это может свидетельствовать об агрессивном поведении между тираннозаврами, но точно неизвестно, были ли тираннозавры активными каннибалами или просто участвовали в межвидовой борьбе (за территорию или право спаривания)[1]. Тем не менее, дальнейшие исследования показали, что повреждения на лицевых костях, в малоберцовой кости и позвонках были получены вследствие инфекционного заболевания, либо окаменелости были повреждены уже после смерти ящера, к тому же травмы эти имеют слишком общий характер, чтобы свидетельствовать о межвидовом конфликте[1].
Некоторые учёные высказались против версии о тираннозавре-падальщике, объясняя это тем, что если тираннозавр действительно являлся таковым, то главным хищником позднемеловой Америки должен был быть другой динозавр. Основными видами добычи были цератопсы, маргиноцефалы и орнитоподы. Другие тираннозавриды имели столько общих черт с тираннозавром, что главными хищниками можно назвать лишь небольших дромеозавров (таких как арчерораптор), троодонтид, крупных вараннобразных ящериц (таких как палеосанива) и недавно обнаруженного дакотараптора, достигавшего более чем 5 метровой длины[1]. Ученые, считающие тираннозавра падальщиком, полагают, что его физическая сила и размеры позволяли ему отбирать добычу у более мелких плотоядных[1]. Однако весьма вероятно, что они с трудом могли найти достаточное количество падали из-за численного превосходства мелких хищников[1] (тираннозавров же было немного, так как они, скорее всего, жили на больших участках площадью примерно в 100 км²[1]). Большинство палеонтологов придерживаются мнения, что тираннозавр был и активным хищником, и падальщиком, как и многие плотоядные больших размеров.
В настоящее время также распространена точка зрения, что тираннозавры, в зависимости от размеров и возраста, занимали разные экологические ниши, подобно современным крокодилам и варанам. Так, новорожденные детеныши скорей всего питались мелкой добычей, по мере роста переходя на все более и более крупную как относительно, так и абсолютно. Не исключено, что самые крупные тираннозавры как раз и промышляли падалью, отнимая добычу у более мелких сородичей[1].
Существует гипотеза, что тираннозавр мог умерщвлять жертву с помощью своей зараженной слюны. Впервые эту теорию предложил Уильям Аблер[1]. Он изучил зазубрины на зубах тираннозавра и сделал вывод, что между ними могли скапливаться сгнившие остатки мяса, вследствие чего тираннозавр обладал смертельным укусом, заражавшим жертву вредными бактериями (ранее считалось, что подобное биологическое оружие использовал комодский варан). Джек Хорнер обратил внимание на то, что зазубрины у тираннозавра имеют скорее кубическую форму, в то время как у комодского дракона они закруглены[1]. Более того, вредные бактерии могут содержаться в слюне у любого животного, поэтому вопрос о том, использовал ли их тираннозавр для умерщвления жертвы, остаётся открытым.
Как и многие другие тероподы, тираннозавр, вероятно, вырывал из туши куски мяса, тряся голову из стороны в сторону в боковом направлении, примерно также, как это делают крокодилы. Голова у тираннозавра была не такой подвижной, как у аллозавроид, из-за плоских сочленений шейных позвонков[1]. Однако, по мнению специалистов, за один укус взрослый тираннозавр мог вырвать из тела жертвы кусок мяса весом в 70 кг[1].
Каннибализм
Исследование, проведенное в 2010 году палеонтологами Карри, Хорнером, Эриксоном и Лонгричем, подняло вопрос о том, был ли каннибализм свойственен тираннозаврам. Они изучили несколько экземпляров тираннозавров с отметинами, оставленными зубами динозавров этого же рода. Подобные отметины были обнаружены в плечевой кости, костях стопы и плюсне. Однако они свидетельствуют не о межвидовых столкновениях, а о том, что тираннозавры могли питаться останками своих сородичей (поскольку в схватке ящерам было тяжело добраться до этих частей тела). То, что отметины были оставлены на наименее мясистых участках туши, означает, что либо мясистые части уже были съедены, либо к моменту обнаружения трупа они уже сгнили. Возможно, что подобное поведением наблюдалось и у других тираннозаврид[1].
Стайные охотники?
Филип Джон Карри, палеонтолог из Университета Альберты, высказал предположение о том, что тираннозавры могли жить в стаях. Карри сравнил тираннозавра рекс с близкородственными ему видами — Tarbosaurus bataar и Albertosaurus sarcophagus, для которых Карри ранее уже нашел окаменелые свидетельства стайной жизни[1]. Карри отмечал, что в Южной Дакоте была сделана находка скелетов трех тираннозавров в непосредственной близости друг от друга[1]. Выполнив компьютерную томографию, Карри заявил, что размер мозга тираннозавра в три раза превышает обычный размер для животного таких габаритов, следовательно, ящер был способен к сложному социальному поведению. В частности, отношение размеров мозга тираннозавра к общему размеру тела превышало этот показатель у крокодилов и было в три раза выше, чем у таких травоядных, как трицератопс. По мнению Карри, тираннозавр был в шесть раз умнее большинства других динозавров и рептилий[1][1]. Карри объяснял необходимость стайной охоты тем, что жертвы тираннозавров были либо хорошо вооружены (трицератопс и анкилозавр), либо могли быстро бегать. Учёный также отмечал, что ювенильные и взрослые особи охотились вместе: небольшие и, следовательно, проворные молодые особи преследовали жертву, а взрослые использовали свою силу для её умерщвления (эта стратегия охоты наблюдается и у современных стайных хищников)[1].
Тем не менее, гипотеза Карри о стайной охоте тираннозавров подверглась жесткой критике со стороны других специалистов. Брайан Суитек в своей статье для газеты The Guardian от 2011 года[1] отмечал, что гипотеза Карри не была представлена в реферируемых научных журналах, а лишь в телевизионном выпуске и написанной по нему книге «Dino Gangs» («Банды динозавров»). Помимо этого, Суитек заявил, что Карри основывается лишь на свидетельствах стайной жизни у современных хищников, а также другого вида динозавров (Tarbosaurus bataar), к тому же доказательства подобного образа жизни у T. bataar тоже не были опубликованы и представлены научному сообществу. Согласно Суитеку и некоторым другим ученым, вступившим в дискуссии о телевизионном выпуске «Dino Gangs», доказательства стайной жизни для тираннозавра и альбертозавра весьма неубедительны, так как основываются лишь на найденных рядом скелетах, которые могли оказаться вместе вследствие наводнений или прочих бедствий. По мнению Суитека, месторождение костей альбертозавра, на котором основывался Карри, как раз предоставляет геологические свидетельства наводнения. Суитек писал:
Шаблон:Начало цитаты
…поведение динозавров невозможно воссоздать, основываясь исключительно на костях. Геологический контекст, в котором были обнаружены кости <…> необходим для исследования жизни и смерти динозавров
Шаблон:Конец цитаты
Шаблон:Oq
[1], а также отмечал, что прежде, чем делать выводы о социальном поведении, Карри должен описать окружающую среду, какой она была на месте захоронения останков тираннозавров. Суитек назвал те сенсационные заявления, которые делала пресса и новости о программе «Dino Gangs», «тошнотворным надувательством», и обвинял компанию Atlantic Productions, транслировавшую программу, в постоянных мистификациях, вспоминая уже имевший место скандал с Darwinius masillae[1].
О своём мнении насчет теории Карри высказывался также Лоренс Уитмер. Он писал, что социальное поведение не может быть воссоздано по эндокранам мозга, так как даже у леопардов, которые ведут одинокий образ жизни, мозг выглядит так же, как у львов, живущих и охотящихся в группах. Уитмер заявил, что максимум, что могут сделать специалисты — изучить размеры конечного мозга тираннозавра, сравнить с современными животными и сделать некоторые измерения. По мнению Уитмера, тираннозавры обладали достаточно большим мозгом, чтобы охотиться сообща. Учёный полагает, что совместной охотой (Шаблон:Lang-en) можно назвать тот процесс, при котором каждый хищник атакует ту или иную особь травоядного животного, а под согласованной охотой (Шаблон:Lang-en) он подразумевает нечто более сложное. Также он придерживается мнения, что в процессе эволюции хищники перешли от охоты в одиночку к согласованной охоте, причем совместная охота, которой занимались тираннозавры, является неким «промежуточным звеном». Специалист заявил, что главной особенностью согласованной охоты является то, что член стаи в случае своего индивидуального успеха не пожирает жертву сразу после её умерщвления, а продолжает охоту вместе со своими сородичами (например, гонит жертву по направлению к другим хищникам), зная, что при таком раскладе охота может оказаться более результативной. Также Уитмер говорил о том, что не может однозначно утверждать, были ли тираннозавры способны к согласованной охоте, так как сведений, которые предоставляют эндокраны, недостаточно. Согласованная охота увеличила бы результативность, но в то же время повысила бы степень риска[1].
23 июля 2014 года в Канаде были впервые найдены окаменевшие Шаблон:Нп3, которые могут говорить в пользу того, что тираннозавры могли охотиться стаями[1][1].
Палеопатология
В 2001 году Брюс Ротшильд со своими коллегами опубликовал исследование, рассмотревшее окаменелые свидетельства Шаблон:Нп3 и отрывных переломов у теропод, а также последствий поведения этих ящеров. В отличие от других травм, усталостные переломы были вызваны постоянными нагрузками на конечности, а не какими-либо неожиданными происшествиями. Всего исследователи изучили 81 кость стопы тираннозавров, и только у одной из них был обнаружен усталостный перелом, и ни у одной из 10 костей передних конечностей такого повреждения не было. Палеонтологам удавалось находить отрывные переломы только у тираннозавров и аллозавров. В частности, отрывной перелом обнаружили на плечевой кости тираннозавра «Сью», на том участке, где располагались дельтовидная и большая круглая мышцы. Тот факт, что и у тираннозавров, и у аллозавров отрывные переломы находили только в пределах передних конечностей и плечевого пояса, говорит о том, что у тероподов была более сложная мускулатура, чем у птиц, и имела функциональные отличия. Учёные полагают, что разрыв сухожилия у тираннозавра «Сью» был вызван тем, что она пыталась схватить активно сопротивлявшуюся жертву. Кроме того, наличие подобных переломов указывает на то, что тираннозавры скорее вели образ жизни активных хищников, чем падальщиков[1].
Исследование 2009 года показало, что дыры, обнаруженные в черепе у некоторых экземпляров тираннозавров, которые прежде считали ранами, полученными в межвидовых конфликтах, на самом деле были вызваны паразитами вроде трихомонады — простейшего, которое обычно поражает птиц[1]. Эти паразиты способны не только поражать мягкие ткани, но и делать в костях рубцы изнутри. Возможно, паразиты передавались через воду, которую пили зараженные динозавры[1].
Свидетельства в пользу межвидовых столкновений были найдены Джозефом Питерсоном и его коллегами у ювенильной особи тираннозавра, которую они окрестили «Джейн». Питерсон со своими сотрудниками обнаружил впоследствии зажившие колотые раны на нижней челюсти и морде и предположил, что их нанес другой молодой тираннозавр. Последующая компьютерная томография подтвердила гипотезу Питерсона[1]. Специалисты также обратили внимание на то, что повреждения у «Джейн» отличались по структуре от тех, что были найдены у «Сью» и располагались на других участках[1].
Останки тираннозавра были найдены в отложениях лансского яруса, жил ящер в эпоху позднего мелового периода, в маастрихтский век. Ареал тираннозавра простирался от Канады до штатов Техас и Нью-Мексико, США. В северных районах ареала среди травоядных доминировал трицератопс, а в южных — зауропод аламозавр. Останки тираннозавров были найдены в различных экосистемах — удаленных от моря участках суши, болотистых местностях, засушливых и полузасушливых (аридных и полуаридных) равнинах. Несколько примечательных находок тираннозавра было сделано в формации Хелл-Крик. В эпоху маастрихтского века этот район был субтропическим, с теплым и влажным климатом. Флора была представлена преимущественно цветковыми растениями, но также встречались хвойные деревья вроде метасеквои и араукарии. Тираннозавр делил среду обитания с трицератопсом, а также близкородственным трицератопсу торозавром, утконосым эдмонтозавром, закованным в панцирь анкилозавром, пахицефалозавром, представителем гипсилофодонтид тесцелозавром и тероподами орнитомимом и троодоном[1]
Другое месторождение останков тираннозавра — Шаблон:Нп3 Вайоминга. Считается, что миллионы лет назад она представляла собой байу — экосистему, похожую на современное побережье Мексиканского залива. Фауна этой формации очень схожа с фауной Хелл-Крик, но нишу орнитомима занимал струтиомим, кроме того, там обитал небольшой представитель цератопсов под названием Шаблон:Нп3[1].
В южных районах своего ареала тираннозавр жил бок о бок с аламозавром, торозавром, эдмонтозавром, возможно, с представителем анкилозавров Шаблон:Нп3 и гигантским птерозавром кетцалькоатлем[1]. В тех местах преобладали полузасушливые равнины, на месте которых ранее располагалось Западное внутреннее море[1].
История изучения
Генри Файрфилд Осборн, президент Американского музея естественной истории, придумал название Tyrannosaurus rex в 1905 году. Оно переводится как «королевский ящер-тиран», тем самым Осборн подчеркивал размеры животного и доминирование над другими животными того времени[1].
Ранние находки
Зубы, сейчас приписываемые тираннозавру рекс, нашел в 1874 году Артур Лейкс около Голдена, штат Колорадо. В начале 1890-х Джон Белл Хэтчер коллекционировал посткраниальные элементы скелета, найденные на востоке Вайоминга. Тогда считалось, что эти окаменелости принадлежат крупной особи орнитомима, но сейчас их приписывают тираннозавру. А в 1982 году Эдвард Дринкер Коп нашел в Южной Дакоте фрагменты позвоночника и описал новый вид динозавра — Manospondylus gigas[1].
Барнум Браун, помощник куратора Американского музея естественной истории, нашел первый частично сохранившийся скелет тираннозавра в 1900 году в восточном Вайоминге и предоставил останки Осборну, который по ним в публикации от 1905-го описал новый вид — Dynamosaurus imperiosus. В 1902 году Браун нашел другой частичный скелет в формации Хелл-Крик, Монтана, и в той же публикации, в которой был описан D. imperiosus, Осборн приписал эти останки новому виду, который он назвал Tyrannosaurus rex[1]. В 1906-м Осборн признал D. imperiosus синонимом T. rex, а последнее — валидным названием вида[1]. Теперь окаменелости, приписанные D. imperiosus, хранятся в Лондонском музее естествознания[1].
Общее количество частичных скелетов тираннозавра, найденных Брауном, равняется пяти. В 1941 году находка Брауна от 1902-го была продана Музею естественной истории Карнеги, Питтсбург, штат Пенсильвания. Четвёртая, самая большая находка Брауна, сделанная в формации Хелл-Крик, была выставлена в Американском Музее естественной истории в Нью-Йорке[1].
Несмотря на то, что скелетов тираннозавра было найдено множество, палеонтологам до сих пор удалось обнаружить только одну цепочку окаменевших следов — у Ранчо Филмонт-Скаут на северо-востоке Нью-Мексико. Она была обнаружена в 1983 году, а идентифицирована и представлена научному сообществу в 1994-м[1].
Маноспондил или тираннозавр рекс?
Первый названный экземпляр, который можно приписать тираннозавру рексу, состоит из двух частично сохранившихся позвонков (один из которых утерян), найденных Эдвардом Копом в 1892 году. Коп считал, что они принадлежат представителю цератопсид, и назвал их «маноспондилом» (Шаблон:Lang-lat — «гигантский пористый позвонок», в связи с многочисленными отверстиями для кровеносных сосудов, найденными в окаменелостях)[1]. Позднее ученые пришли к выводу, что эти останки принадлежали скорее тероподу, чем цератопсиду, и в 1917 году Осборн заметил схожие черты в останках M. gigas и T. rex. Тем не менее тогда он не мог синонимизировать эти два вида, поскольку в его руках были лишь останки фрагментарного характера[1].
В июне 2000 года геологический институт Шаблон:Нп3 отыскал местонахождение останков M. gigas и раскопал там ещё некоторые останки тираннозавров, которые принадлежали этому же виду и были идентичны останкам вида Tyrannosaurus rex[1]. Согласно правилам Международного кодекса зоологической номенклатуры, название Manospondylus gigas должно было быть в приоритете над именем Tyrannosaurus rex, поскольку было дано ранее. Тем не менее, в четвёртом издании кодекса, вышедшем 1 января 2000 года, говорится, что «должно быть принято название, которое имеет преобладающее применение» в случае, если «ранний синоним или омоним не использовался как законное наименование после 1899 года» и если «поздний синоним или омоним использовался для отдельного таксона и был упомянут не менее чем в 50 работах, изданных не менее чем 10 авторами в последующие 50 лет…»[1] Согласно этим условиям, имя Tyrannosaurus rex является законным и может считаться «защищенным названием» (Шаблон:Lang-lat), в то время как Manospondylus gigas можно считать «забытым названием» (Шаблон:Lang-lat)[1].
В культуре
Со времени своего открытия в 1905 году тираннозавр стал одним из наиболее популярных видов динозавров. Это единственный динозавр, который наиболее известен широкой публике по своему видовому, а не родовому названию (тираннозавр рекс), учёные также часто используют сокращение T. rex[1]. Роберт Бэккер отмечает эту особенность в своей книге Шаблон:Нп3. По его мнению, видовое название является более звучным[1].
Одно из первых появлений тираннозавра на киноэкране произошло в классическом голливудском фильме «Кинг-Конг» 1933 года. Появляется он практически и во всех экранизациях романа Артура Конана Дойля «Затерянный мир».
Пожалуй, наиболее знаменитым появлением тираннозавра в кино является фильм Стивена Спилберга «Парк юрского периода» (1993), и далее в его сиквелах — «Парк юрского периода 2: Затерянный мир» (1997) и «Парк юрского периода 3». В первых двух фильмах он был показан в качестве главного злодея, в третьей части появляется эпизодически в схватке со спинозавром. Тираннозавр также присутствует в финале фильма «Мир юрского периода», где сражается с Индоминусом Рексом и наносит ему значительные повреждения. В нетрадиционном образе практически домашнего питомца, а не свирепого хищника и убийцы, тираннозавр (вернее, его скелет) был показан в фантастической комедии 2006 года «Ночь в музее».
Тираннозавр рекс также является главным героем шестой серии («Гибель династии») научно-популярного сериала «Прогулки с динозаврами»[1].
Наряду с этим он также присутствует в ряде других художественных фильмов: «Т-Рекс: Исчезновение динозавров», «Теодор Рекс», «Тэмми и Ти-Рекс».
Образ тираннозавра присутствует и в мультфильмах. Под именем «Острозуб» тираннозавр фигурирует в популярной серии американских полнометражных мультфильмов «Земля до начала времён». Юный тираннозавр по имени Бадди — один из героев мультфильма «Поезд динозавров». Игрушечный тираннозавр Рекс — персонаж серии мультфильмов «История игрушек».
Не обошли тираннозавра и писатели. Тираннозавр является главным объектом охоты героев рассказа Л. Спрэг де Кампа Шаблон:Нп3.
В честь тираннозавра также была названа английская рок-группа T. Rex.
Примечания
Шаблон:Примечания
Литература
| Язык
| {{{Язык}}}
|
| {{{Название}}}
|
[[Изображение:{{{Обложка}}}|250px]]
|
| Под редакцией
| {{{Редакторы}}}
|
| Авторы
| {{{Авторы}}}
|
| Город издания
| {{{Город}}}
|
| Издательство
| {{{Издательство}}}
|
| Год издания
| {{{Год}}}
|
| Количество страниц
| {{{Страницы}}}
|
| Издание
| {{{Издание}}}
|
| ISBN
| {{{ISBN}}}
|
| УДК
| {{{УДК}}}
|
| doi
| {{{doi}}}
|
| ББК
| {{{ББК}}}
|
| Тираж
| {{{Тираж}}}
|
| Формат
| {{{Формат}}}
|
| Печатные листы
| {{{ПечатЛист}}}
|
| Гарнитура
| {{{Гарнитура}}}
|
| Подписано в печать
| {{{ВПечать}}}
|
| Художник
| {{{Художник}}}
|
| Корректор
| {{{Корректор}}}
|
Ссылки






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}